
¿Nuestro cerebro activa las mismas redes neuronales cuando tiene que tomar una decisión con riesgo que cuando debe hacer una elección segura? Veámoslo con un ejemplo. Imaginemos que nos encontramos en una exposición sobre turismo y ocio. En uno de los stands, unos desconocidos están repartiendo cajas. Nada más aproximarnos, nos dan la nuestra. No es muy grande, pero enseguida vemos de qué se trata. En el interior de esa caja de plástico transparente vemos el libro El Principito, de Antoine de Saint-Exupéry, junto a una nota escrita a mano donde se lee: «Es el tiempo que te dedicas lo que te hace tan importante». Ahora que sabemos cuál es el contenido de la caja, si decidimos quedárnosla, tendremos un ejemplar de El Principito en casa, aparte de un recuerdo positivo que asociaremos con Francia (país propietario del stand); si por el contrario devolvemos la caja a esos desconocidos, nos iremos con las manos vacías. Es una decisión sencilla, pero lo más importante es que no implica riesgo alguno, pues sabemos con total certeza qué ocurrirá en función de la decisión que tomemos.
Sin embargo, avanzando un poco más por el laberinto de pasillos que forman los distintos expositores y antes de entrar en el último pabellón, llegamos a otro stand donde tenemos la posibilidad de elegir entre dos cajas (una amarilla y una verde). Una contiene un viaje con los gastos de vuelo y hotel pagados a un destino que solo sabremos dos días antes de la fecha de partida, mientras que la otra, en cambio, puede ser cualquier cosa, e incluso nada. Desconocemos qué caja contiene cada cosa. ¿Nos animaríamos a participar y elegir una caja? De ser así, ¿con cuál nos quedaríamos, con la amarilla o con la verde?
¿El análisis que hemos hecho para tomar estas dos decisiones ha sido el mismo en ambos casos? Lógicamente, no. Lo más probable es que, en el primer caso, nos hayamos preguntado si ya tenemos ese libro en casa para devolver la caja a los desconocidos, o hayamos pensado sencillamente «a caballo regalado no le mires el diente» y hayamos aceptado la caja. En cambio, en el segundo caso, posiblemente hemos tenido que valorar si nos gusta viajar sea cual sea el destino, pensar si podremos irnos de viaje en las fechas que nos indiquen, preguntarnos qué premio quizá ganemos si no escogemos la caja con el viaje, o calcular cómo nos sentiremos si realmente la caja está vacía, por nombrar algunas posibilidades.
Pues bien, si el análisis no ha sido el mismo en estas dos situaciones, una sin riesgo y otra con riesgo, ¿por qué nuestro cerebro debería activar las mismas áreas corticales y subcorticales al tomar la decisión en cada uno de los dos casos?
Para poder responder a esta pregunta nos gustaría comenzar hablando de qué sucede en nuestro cerebro cuando tomamos decisiones carentes de incertidumbre, pues muchos artículos científicos apuntan a que hay procesos neuronales distintos cuando se trata de tomar una decisión con riesgo frente a otra que no lo tiene. En inglés dichos procesos se conocen con el nombre de hot process y cold process, respectivamente.
En 2008, Andrew Lawrence, Luke Clark, Jamie Labuzetta y Barbara Sahakian publicaron «The innovative brain» como resultado del estudio que realizaron sobre el cerebro del emprendedor. Para analizar las decisiones sin riesgo (cold) utilizaron un juego conocido como Torre de Londres (véase la figura 13), donde simplemente hay que ir moviendo las distintas bolas de colores, de una en una, hasta colocarlas en el orden que te han indicado al inicio de la partida. Dicho juego requiere planificación, memoria y capacidad de decisión. Los investigadores encontraron que este tipo de decisiones, emocionalmente neutras sin ponderación alguna de recompensa o castigo, produce activación en la corteza prefrontal dorsolateral, encargada de planificar y controlar el comportamiento con el fin de alcanzar el objetivo deseado. No obstante, en 2009, Sharlene Newman, John Greco y Donghoon Lee, sirviéndose de la misma prueba, añadieron al grupo de áreas cerebrales que participan en este tipo de decisión a la corteza prefrontal derecha, sobre todo las regiones asociadas con procesos ejecutivos como la memoria de trabajo; la corteza parietal, relacionada con el procesamiento visual-espacial; los ganglios basales, implicados en aspectos motores y en funciones más ejecutivas necesarias para resolver la tarea, y la corteza cingulada anterior, cuya principal función es la detección de errores y el control de conflictos en tareas de percepción.
Entonces, ¿estas áreas cerebrales únicamente participan en la toma de decisiones sin riesgo? Y si fuese así, ¿qué ocurre cuando existe incertidumbre o riesgo?
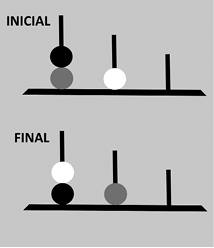
| Figura 13: Torre de Londres. Se trata de una prueba neuropsicológica que consiste en reorganizar las bolas para pasar de la situación inicial a la final con el menor número posible de movimientos, teniendo en cuenta que solo se puede mover una bola cada vez. Se utiliza para evaluar la capacidad de planificación y ejecución de las personas. |
El riesgo es, desde hace mucho tiempo, un tema que interesa profundamente a la comunidad científica. Gracias a experimentos con animales y humanos se ha avanzado en el estudio de la toma de decisiones con riesgo, evaluando aspectos como la recompensa o la valoración de las opciones, ya que estos van a condicionar de manera significativa nuestra decisión de arriesgarnos o, por el contrario, de permanecer en la certeza, en la seguridad. Por ejemplo, el circuito neuronal de la recompensa está formado por áreas dopaminérgicas del cerebro medio y de los ganglios basales que se proyectan a estructuras estriadas, como el estriado ventral y dorsal, así como al lóbulo frontal, concretamente a la corteza prefrontal ventromedial. Se ha comprobado que hay actividad en estas áreas dopaminérgicas con las recompensas actuales y las anticipadas.
Una de las áreas con más capacidad dopaminérgica es el núcleo accumbens, considerado el centro neuronal de la recompensa. De hecho, el proceso de razonamiento y conducta comienza con modulaciones y fluctuaciones de la dopamina. Este proceso neurobiológico viene determinado por el aprendizaje de estas neuronas dopaminérgicas, que en un principio dan respuesta a la recompensa real, pero que poco a poco son capaces de asociar dicha recompensa con otros estímulos sociales, de tal forma que se activan con anticipación. Schultz las llamó «neuronas de predicción», ya que estaban más pendientes de la predicción de las recompensas que de las recompensas propiamente dichas.
Las neuronas dopaminérgicas pueden responder a diferentes estímulos asociados que preceden a la recompensa real, como una melodía, una luz, etcétera, de tal forma que si la predicción es correcta y coincide con la recompensa real, la neurona aumenta la tasa de disparo de dopamina. Es decir, las neuronas dopaminérgicas aprenden constantemente pautas basadas en la experiencia y comparan estas predicciones con lo que sucede en la realidad, con lo que crean modelos de correlación que permiten al cerebro anticipar lo que sucederá después. En este sentido, entendemos que el ser humano muestra diferencias considerables en la modulación de la dopamina, en la medida en que su comportamiento refleja un sesgo hacia la búsqueda de gratificación o la evitación de los estímulos aversivos, lo cual da lugar a diferencias individuales según sea el aprendizaje por recompensa o por castigo. Es el caso de aquellas personas que buscan la novedad o el riesgo.
La corteza orbitofrontal también participa en este circuito, gracias a su capacidad de control, evaluación y ejecución de la conducta ante las diferentes opciones que se le presentan antes de tomar la decisión. Los pacientes con lesiones en esta área tienden, en el ámbito de la economía, a perder cuantiosas cantidades de dinero, solicitar préstamos constantemente y declararse en quiebra, pues, como consecuencia de un déficit en la modulación dopaminérgica, no son capaces de asociar sus decisiones con sentimientos negativos y deciden de manera aleatoria.
Otra área implicada es la corteza cingulada anterior, que recibe muchas neuronas aferentes dopaminérgicas y que es la responsable final del MS y su integración con la respuesta cognitiva responsable de la toma de decisión (una cuestión explicada ya anteriormente). Esta área ayuda a controlar la «conversación» entre lo que sabemos y lo que sentimos, ajustando las expectativas a la realidad y aprendiendo de las pautas esperadas, es decir, anticipándose al evento. Se encuentra entre el lóbulo frontal y la amígdala. Normalmente la corteza cingulada anterior tiene una función específica de evaluación y detección del «conflicto», e indica la necesidad de implementar procesos estratégicos para reducir el problema y mantener el rendimiento.
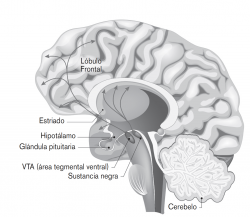
Figura 14: Circuitos dopaminérgicos presentes en la regulación de las emociones y del comportamiento humano.
En la regulación de las emociones y del comportamiento humano hay tres circuitos dopaminérgicos que se organizan de formas diferentes (véase la figura 14):
- Circuito nigroestriado: proyecta sus neuronas dopaminérgicas a los núcleos estriados, principalmente al núcleo caudado y el putamen, y es el encargado de llevar a cabo el control de los procesos motores del comportamiento y de la motivación para iniciar el movimiento.
- Circuito mesolímbico: proyecta sus neuronas dopaminérgicas principalmente al núcleo accumbens y otras estructuras límbicas, entre ellas la amígdala, el hipocampo y el septum, y está implicado en el procesamiento y reforzamiento de estímulos agradables y de recompensa, la experiencia subjetiva del placer y la motivación.
- Circuito mesocortical: proyecta sus neuronas fundamentalmente a la corteza orbito-frontal, la corteza cingulada anterior y la corteza entorinal, y participa en funciones cognitivas tales como la concentración, la memoria de trabajo, el juicio, la planificación y la ejecución de respuestas conductuales.
En conclusión, las emociones humanas se relacionan con las predicciones de unas neuronas altamente sensibles y plásticas, que están constantemente ajustando sus conexiones a la realidad y que cada vez que se comete un error se amoldan a los cambios. Nuestras emociones son respuestas empíricas al medio, por lo que cabría decir que «si me engañas una vez la culpa es tuya, pero si me engañas dos veces la culpa es de mis neuronas dopaminérgicas», que no se han adaptado correctamente al medio.
Siguiendo con la recompensa, muchos autores apuntan a una convergencia en los circuitos neuronales a la hora de valorar diferentes tipos de recompensa, a pesar de la importancia que tienen determinadas áreas cerebrales dependiendo de la clase de recompensa de que se trate. La teoría económica habla del valor subjetivo de los diferentes tipos de recompensa cuando se refiere a estos puntos de convergencia.
En 2011, Dino Levy y Paul Glimcher analizaron esta hipótesis. Para ello examinaron las decisiones arriesgadas que los participantes en el estudio, privados de alimento y agua, tomaron sobre dinero, alimentos y agua. Encontraron que las preferencias de riesgo manifestadas acerca de los distintos tipos de recompensa estaban altamente correlacionadas, es decir, el grado de aversión al riesgo mostrado en las decisiones referentes al dinero era similar al de las elecciones sobre alimentos y agua. A nivel neuronal, como área común de representación del valor subjetivo de la recompensa de dinero y alimentos, encontraron activación en la corteza prefrontal ventromedial. Muchos estudios sustentan que esta área, junto con la corteza orbitofrontal, tiene un papel en la representación del valor en las decisiones arriesgadas y permite comparar recompensas pertenecientes a distintos campos. A nivel particular, en los alimentos fue la región hipotalámica dorsal, mientras que en el dinero fue la corteza cingulada posterior. Otras investigaciones han relacionado la corteza cingulada anterior y posterior con la incertidumbre y la ambigüedad al tomar una decisión. Levy y Glimcher sugieren que las distintas redes de valoración de las recompensas convergen en una red de valoración unificada, lo cual permite la comparación directa de varias recompensas para guiar la toma de la decisión.
Otros autores, como Elke Weber, Ann-Renee Blais y Nancy Betz (2002), desde el punto de vista de las teorías psicológicas, consideran que la aversión al riesgo que muestra una persona no es independiente del tipo de decisión que esté tomando, es decir, que un individuo sea arriesgado en el ámbito de las finanzas no implica necesariamente que también lo sea en aspectos relacionados con la salud o el ocio, como apuntan Levy y Glimcher. Weber, Blais y Betz argumentan que resolvemos de formas diferentes las cuestiones, tanto personales como laborales, que implican un determinado nivel de riesgo e incertidumbre. Por este motivo desarrollaron una escala psicométrica para medir el riesgo conocida como DOSPERT (Domain-Specific Risk-Taking), cuya última versión, realizada en 2006, consta de treinta preguntas relacionadas con los campos de las finanzas, la salud y el ocio y con aspectos sociales y éticos. A grandes rasgos, y según apuntan estos autores, se podría decir que es como si, para cada tipo de decisión, el «marcador» de aversión al riesgo empezase a contar desde cero. Sin embargo, estos investigadores sí confirman una correlación en las preferencias entre los distintos tipos de recompensas.
Un año más tarde, en 2012, Levy y Glimcher, planteándose la pregunta «¿Cómo decidimos en función del tipo de recompensa?», revisaron trece estudios llevados a cabo con resonancia magnética funcional y corroboraron la conclusión de la «red neuronal común» para la valoración subjetiva de los diferentes tipos de recompensas. De modo que existe una «representación común» de la recompensa localizada en una subregión de la corteza prefrontal ventromedial, al lado de la corteza orbitofrontal medial, lo cual sugieren que ayuda a lograr una fundamentación neurobiológica de la teoría económica, así como una herramienta para medir las preferencias.
Para ilustrar su propuesta, Levy y Glimcher elaboraron el siguiente esquema, en el que muestran las redes neuronales que intervienen en la toma de la decisión (véase la figura 15). Sugieren que la información procedente de las estructuras corticales y subcorticales converge hacia una única representación del valor común antes de pasar a las áreas motoras previas a la elección. Los procesos de percepción del medio ambiente se dirigen desde las estructuras subcorticales, en las que se modulan los procesos de arousal, esto es, de nivel de activación cerebral, hasta llegar a las áreas corticales responsables del proceso de representación común de los mismos antes de llevar a cabo la respuesta motora.
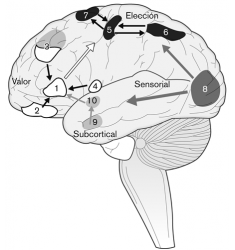
Figura 15: Redes neuronales que intervienen en la toma de decisiones. Las áreas cerebrales numeradas son: (1) corteza prefrontal ventromedial, (2) corteza orbitofrontal, (3) corteza prefrontal dorsolateral, (4) ínsula, (5) corteza motora primaria, (6) corteza parietal posterior, (7) campos del movimiento ocular, (8) corteza visual, (9) amígdala y (10) estriado. Fuente: Levy y Glimcher, «The root of all value: a neural common currency for choice», 2012
La literatura científica también apoya la idea de que la corteza prefrontal ventromedial y la corteza orbitofrontal medial, con la información transmitida por el estriado y el resto de la red neuronal encargada de procesar la recompensa, responden a las ganancias, a las pérdidas y a sus respectivas probabilidades. La menor activación de estas áreas se ha relacionado con la anticipación de pérdidas, así como con la disminución del valor de la opción, por ejemplo, cuando las probabilidades de obtener ganancias son más bajas o se prevén demoras en la recepción de las recompensas. Hablamos del valor de la opción porque cuando tomamos decisiones económicas, para poder elegir, damos valores a las distintas opciones. Estas dos regiones, la corteza prefrontal ventromedial y la corteza orbitofrontal, son las responsables de la representación del valor subjetivo en nuestro cerebro.
Se cree que la corteza orbitofrontal interviene en el cálculo de la recompensa determinando su valor en función de lo gratificante que sea la recompensa. Y esta información se transmite a la corteza prefrontal dorsolateral, la cual se va a encargar de controlar el comportamiento con el fin de lograr el objetivo deseado.
Otro binomio de áreas cerebrales que hay que destacar es el de la corteza cingulada anterior y la corteza prefrontal dorsomedial. Diversos experimentos asignan a estas áreas la función de resolver el conflicto cognitivo o decisional frente a opciones similares. La activación en estas áreas aumenta cuando la diferencia entre el valor de cada opción disminuye. En un estudio realizado en 2008, Jean-Baptiste Pochon, Jason Riis, Alan Sanfey, Leigh Nystrom y Jonathan Cohen encontraron que la actividad en la corteza cingulada anterior y la corteza prefrontal dorsomedial era mayor cuando los voluntarios que participaron en el estudio tenían un conflicto porque debían escoger una de varias caras atractivas muy parecidas. Asimismo, sugirieron que el papel de la corteza cingulada anterior, en tareas de decisión complejas, es analizar el conflicto en la fase de decisión.
Un aspecto importante de la toma de decisiones con riesgo que influye en la valoración y la recompensa son las probabilidades que tenemos para cada opción. En relación con la conducta, las probabilidades de riesgo también pueden llegar a condicionar nuestras decisiones. Es aquí donde la psicología destaca el importante papel que juega la actitud de las personas frente al riesgo, ya que el nivel de tolerancia de la incertidumbre difiere en cada individuo. Volviendo a la neurociencia, algunos estudios apuntan a la implicación de la corteza prefrontal medial y la corteza parietal cuando conocemos las probabilidades asociadas a cada uno de los resultados. Sin embargo, es sobre todo la ínsula anterior la que se activa cuando decidimos sin saber cuáles pueden ser las consecuencias, así como el caudado, asociado a la motivación ejecutoria.
En 2005, Ming Hsu, Meghana Bhatt, Ralph Adolphs, Daniel Tranel y Colin Camerer publicaron un artículo titulado «Neural systems responding to degrees of uncertainty in human decision-making» que trata del circuito neuronal que subyace en los procesos de toma de decisiones en ambientes de riesgo (probabilidades conocidas) y ambigüedad (probabilidades desconocidas).
Desde el punto de vista de las ciencias sociales, las únicas variables que deberían influir en la toma de decisiones con incertidumbre son las probabilidades y los posibles resultados. La teoría de la utilidad esperada está de acuerdo con eso, pero matiza que la confianza que uno tenga en esas probabilidades no debería afectar a la elección. Sin embargo, numerosos estudios han observado que preferimos apostar por el riesgo frente a la ambigüedad, es decir, conocer o desconocer las probabilidades sí nos afecta a la hora de tomar una decisión, como ya apuntaba Ellsberg cuando formuló su paradoja. Este fenómeno se conoce con el nombre de «aversión a la ambigüedad».
En cambio, hay otros autores que no se centran en las probabilidades de manera particular, sino que destacan que la decisión, de lo que realmente depende es de cuánta información relevante falta o de lo ignorantes que se sienten las personas en comparación con otros.
Basándose en lo que dice la neurociencia sobre recompensa e incertidumbre, Hsu, Bhatt, Adolphs, Tranel y Camerer llevaron a cabo un experimento, dividido en tres partes, en el que jugaban con la cantidad de información desconocida por el voluntario durante la prueba.
Encontraron que el nivel de ambigüedad estaba correlacionado positivamente con la activación de la amígdala (reacciones emocionales) y la corteza orbitofrontal (integración de la información cognitiva y emocional). Estas áreas cerebrales reciben información sensorial multimodal y trabajan de manera conjunta analizando el valor de los estímulos, incluidos aquellos cuyo valor es incierto. Pero también descubrieron una correlación negativa con el sistema estriado (valor esperado de las decisiones), pues la actividad del estriado está positivamente correlacionada con la recompensa esperada. En otras palabras, a medida que aumenta la ambigüedad, se incrementa la actividad de la amígdala y la corteza orbitofrontal y decrece la del estriado, mientras que cuando mejora la recompensa, se intensifica la actividad del estriado. Las principales áreas cerebrales que se activan cuando una persona tiene que tomar una decisión con ambigüedad son la corteza orbitofrontal, la amígdala y la corteza prefrontal dorsomedial. En definitiva, estos investigadores afirman que, en condiciones de incertidumbre, el cerebro se da cuenta de que falta información y moviliza recursos cognitivos para buscar más datos con los que poder tomar una decisión.
Pero ¿cómo se procesa el riesgo?
A pesar de estos y otros avances, todavía se sabe poco sobre cómo el cerebro procesa el riesgo. Esto se debe, principalmente, al papel que desempeña la emoción en este tipo de decisiones, pues no se sabe con certeza de qué manera participan las emociones cuando nuestro cerebro está procesando el riesgo mientras sopesa las potenciales pérdidas. En 2010, Peter Mohr, Guido Biele y Hauke Heekeren, tras revisar unos experimentos realizados con resonancia magnética funcional, llegaron a la conclusión de que la ínsula anterior participa activamente en la representación del riesgo y la visualización de las pérdidas potenciales, posiblemente debido a que interviene en el procesamiento de emociones negativas como la ansiedad, la decepción o el arrepentimiento. Esto sugiere que las pérdidas potenciales también modulan el procesamiento del riesgo. Por otra parte, cabe destacar la actividad detectada en la corteza prefrontal dorsolateral y la corteza parietal en situaciones de riesgo en las que tenemos la posibilidad de decidir, que no se detecta cuando no disponemos de esa opción. Otros investigadores han resaltado el papel que desempeña la corteza prefrontal dorsolateral en el control cognitivo de la emoción durante la toma de decisiones con riesgo, mediante la inhibición o reevaluación de respuestas, recompensas y emociones, lo cual significa que una mayor activación de esta área se relaciona con una menor asunción de riesgos.
De acuerdo con todo lo expuesto anteriormente, las principales áreas que participan en la toma de decisiones con y sin riesgo son las que pueden apreciarse en la figura 16.
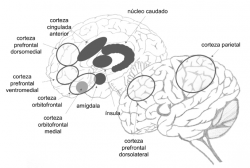
Figura 16: Áreas implicadas en la toma de decisiones con riesgo.
Comprender los mecanismos neuronales que subyacen en los procesos que implican riesgos y recompensas es dar un paso de gigante en el entendimiento de las decisiones irracionales que tomamos en nuestra vida. Puestos a soñar, imaginemos que en el futuro logramos desarrollar un dispositivo tecnológico que nos controle la actividad cerebral en tiempo real y nos avise cuando vayamos a tomar decisiones impulsivas muy arriesgadas a la hora de invertir. Esto podría evitar pérdidas cuantiosas de dinero, de lo cual los inversores se alegrarían y las empresas darían gracias. Para ello no solamente es necesario desarrollar la tecnología adecuada, sino también conocer el funcionamiento del cerebro cuando toma este tipo de decisiones, desde que este percibe el estímulo hasta que se produce el mismo acto de elegir. Por esta razón creemos que las investigaciones, dentro de la neuroeconomía, se van a centrar particularmente en los procesos de toma de decisiones en condiciones de riesgo e incertidumbre.
A lo largo de estas páginas hemos respondido a la pregunta planteada al comienzo del apartado, viendo que las redes cerebrales que activamos cuando tomamos decisiones seguras no son exactamente las mismas que activamos cuando tomamos decisiones que conllevan un riesgo. La clave reside en la emoción, el gran enigma de la neurociencia.
Vea nuestro curso de Neuroeconomia
Diferencias entre riesgo e incertidumbre
- Riesgo: En situaciones de riesgo, las probabilidades de los posibles resultados son conocidas o pueden estimarse con precisión. Por ejemplo, en un juego de azar, sabemos las probabilidades de ganar o perder.
- Incertidumbre: En situaciones de incertidumbre, las probabilidades de los resultados no son claras o son completamente desconocidas. Por ejemplo, invertir en un mercado volátil sin información suficiente.
Regiones cerebrales implicadas en la toma de decisiones
El proceso de toma de decisiones bajo riesgo e incertidumbre involucra varias regiones del cerebro, algunas de las cuales están más orientadas a la evaluación racional, mientras que otras responden a factores emocionales.
1. Corteza prefrontal dorsolateral (CPFDL): Evaluación lógica y control cognitivo
La corteza prefrontal dorsolateral está fuertemente asociada con las decisiones racionales y la evaluación lógica de las opciones. Es responsable de:
- El análisis de los resultados y las probabilidades.
- El control de los impulsos para evitar decisiones precipitadas.
- La integración de información nueva para ajustar los planes y estrategias.
Esta región permite tomar decisiones deliberadas y con mayor control, evaluando cuidadosamente las consecuencias a largo plazo, lo que es especialmente importante en situaciones de incertidumbre.
2. Amígdala: Procesamiento emocional y aversión al riesgo
La amígdala es crucial para el procesamiento emocional, particularmente en situaciones que implican riesgo. Está involucrada en:
- La respuesta emocional ante el riesgo, especialmente el miedo y la ansiedad.
- La aversión al riesgo, es decir, la tendencia a evitar decisiones que puedan conllevar pérdidas, especialmente si el marco es negativo.
En situaciones de incertidumbre, la amígdala puede generar una respuesta emocional intensa, lo que puede llevar a decisiones más conservadoras, aunque no necesariamente racionales.
3. Corteza prefrontal ventromedial (CPFVM): Valoración y toma de decisiones
La corteza prefrontal ventromedial está implicada en la valoración de los resultados potenciales de una decisión. Sus funciones incluyen:
- Evaluar el valor subjetivo de una opción, es decir, cómo percibimos personalmente la importancia de una recompensa o pérdida.
- Integrar información emocional y racional para ayudar a tomar decisiones que consideren tanto los beneficios a corto como a largo plazo.
La CPFVM también es esencial en situaciones donde hay que comparar varias alternativas bajo condiciones de incertidumbre, ponderando los riesgos y beneficios de cada una.
4. Estriado (Striatum): Anticipación de la recompensa
El estriado, que incluye el núcleo accumbens, es una región fundamental en el sistema de recompensa del cerebro. Se activa cuando anticipamos una recompensa o un resultado positivo. Sus principales roles incluyen:
- Mediar la motivación para tomar decisiones que se perciben como beneficiosas.
- Evaluar las recompensas inmediatas frente a las recompensas futuras.
En contextos de incertidumbre, el estriado puede influir en la tendencia a asumir riesgos, ya que está vinculado a la búsqueda de recompensas, lo que puede llevar a decisiones más arriesgadas cuando la anticipación de un beneficio es fuerte.
5. Ínsula: Evaluación del riesgo y aversión a la pérdida
La ínsula se activa en situaciones de riesgo y cuando las personas experimentan aversión a la pérdida. Está asociada con:
- La percepción de sensaciones físicas como el malestar, lo que puede llevar a la evitación de riesgos.
- La evaluación de situaciones donde existe una alta probabilidad de pérdida o fracaso.
Cuando enfrentamos incertidumbre o riesgo elevado, la ínsula puede generar respuestas emocionales negativas que aumentan nuestra aversión al riesgo, motivándonos a evitar decisiones que podrían llevar a resultados adversos.
Toma de decisiones bajo riesgo
Las decisiones bajo riesgo, donde las probabilidades de los resultados son conocidas, dependen en gran medida de cómo se percibe el riesgo y el posible beneficio. El cerebro evalúa tanto los costes como las recompensas usando una combinación de regiones responsables del cálculo racional y del procesamiento emocional.
- Evaluación del riesgo:
- La corteza prefrontal dorsolateral trabaja junto a la CPFVM para evaluar los resultados basados en probabilidades, calculando el valor esperado de cada opción.
- Respuestas emocionales:
- La amígdala y la ínsula modulan las respuestas emocionales ante el riesgo, generando emociones como el miedo, lo que puede inclinar la balanza hacia decisiones más conservadoras o evitar riesgos.
- Búsqueda de recompensas:
- El estriado anticipa los posibles beneficios, motivando a las personas a aceptar ciertos riesgos si las recompensas parecen suficientemente atractivas.
Toma de decisiones bajo incertidumbre
Cuando las probabilidades no están claras o no se conocen, las decisiones se complican más y las emociones juegan un papel aún más significativo. El cerebro puede reaccionar de manera más extrema ante la incertidumbre, activando regiones relacionadas con la ansiedad y la aversión al riesgo.
- Mayor incertidumbre, mayor emocionalidad:
- En situaciones de incertidumbre, la amígdala y la ínsula tienden a estar más activas, lo que puede generar mayor temor y una inclinación a evitar tomar decisiones.
- Dificultad para evaluar:
- La corteza prefrontal tiene menos datos concretos con los que trabajar, lo que dificulta el cálculo racional. Esto a menudo lleva a las personas a depender más de heurísticas o atajos mentales, que no siempre son los más precisos.
- Rol de la experiencia:
- En escenarios inciertos, la experiencia previa y la intuición pueden jugar un papel crucial. El cerebro recurre a memorias de situaciones similares para ayudar en la toma de decisiones, utilizando la CPFVM para evaluar si estas experiencias pasadas son útiles en el contexto actual.
Sesgos y toma de decisiones
Los procesos neuronales involucrados en la toma de decisiones bajo riesgo e incertidumbre también están sujetos a sesgosque pueden distorsionar la lógica de la decisión:
- Aversión a la pérdida: Las personas suelen ser más sensibles a las pérdidas que a las ganancias equivalentes. Esto está mediado por la ínsula y la amígdala, que responden más intensamente ante la posibilidad de perder.
- Sesgo de optimismo: El estriado, al anticipar la recompensa, puede llevar a una sobreestimación de los beneficiosen escenarios de riesgo, alentando a asumir riesgos más altos de lo que sería racional.